The Enzyme Commission number (EC number) is a numerical classification scheme for enzymes, based on the chemical reactions they catalyze.[1] As a system of enzyme nomenclature, every EC number is associated with a recommended name for the corresponding enzyme-catalyzed reaction.
EC numbers do not specify enzymes but enzyme-catalyzed reactions. If different enzymes (for instance from different organisms) catalyze the same reaction, then they receive the same EC number.[2] Furthermore, through convergent evolution, completely different protein folds can catalyze an identical reaction (these are sometimes called non-homologous isofunctional enzymes)[3] and therefore would be assigned the same EC number. By contrast, UniProt identifiers uniquely specify a protein by its amino acid sequence.[4]
Format of number[edit]
Every enzyme code consists of the letters «EC» followed by four numbers separated by periods. Those numbers represent a progressively finer classification of the enzyme. Preliminary EC numbers exist and have an ‘n’ as part of the fourth (serial) digit (e.g. EC 3.5.1.n3).[2]
For example, the tripeptide aminopeptidases have the code «EC 3.4.11.4», whose components indicate the following groups of enzymes:
- EC 3 enzymes are hydrolases enzymes (enzymes that use water to break up some other molecule)
- EC 3.4 are hydrolases that act on peptide bonds
- EC 3.4.11 are those hydrolases that cleave off the amino-terminal amino acid from a polypeptide
- EC 3.4.11.4 are those that cleave off the amino-terminal end from a tripeptide
Top level codes[edit]
| Class | Reaction catalyzed | Typical reaction | Enzyme example(s) with trivial name |
|---|---|---|---|
| EC 1 Oxidoreductases | Oxidation/reduction reactions; transfer of H and O atoms or electrons from one substance to another | AH + B → A + BH (reduced) A + O → AO (oxidized) | Dehydrogenase, oxidase |
| EC 2 Transferases | Transfer of a functional group from one substance to another. The group may be methyl-, acyl-, amino- or phosphate group | AB + C → A + BC | Transaminase, kinase |
| EC 3 Hydrolases | Formation of two products from a substrate by hydrolysis | AB + H2O → AOH + BH | Lipase, amylase, peptidase, phosphatase |
| EC 4 Lyases | Non-hydrolytic addition or removal of groups from substrates. C-C, C-N, C-O or C-S bonds may be cleaved | RCOCOOH → RCOH + CO2 or [X-A+B-Y] → [A=B + X-Y] | Decarboxylase |
| EC 5 Isomerases | Intramolecule rearrangement, i.e. isomerization changes within a single molecule | ABC → BCA | Isomerase, mutase |
| EC 6 Ligases | Join together two molecules by synthesis of new C-O, C-S, C-N or C-C bonds with simultaneous breakdown of ATP | X + Y + ATP → XY + ADP + Pi | Synthetase |
| EC 7 Translocases | Catalyse the movement of ions or molecules across membranes or their separation within membranes | Transporter |
Reaction similarity[edit]
Similarity between enzymatic reactions can be calculated by using bond changes, reaction centres or substructure metrics (formerly EC-BLAST], now the EMBL-EBI Enzyme Portal).[6]
History[edit]
Before the development of the EC number system, enzymes were named in an arbitrary fashion, and names like old yellow enzyme and malic enzyme that give little or no clue as to what reaction was catalyzed were in common use. Most of these names have fallen into disuse, though a few, especially proteolyic enzymes with very low specificity, such as pepsin and papain, are still used, as rational classification on the basis of specificity has been very difficult.
By the 1950s the chaos was becoming intolerable, and after Hoffman-Ostenhof[7] and Dixon and Webb[8] had proposed somewhat similar schemes for classifying enzyme-catalyzed reactions, the International Congress of Biochemistry in Brussels set up the Commission on Enzymes under the chairmanship of Malcolm Dixon in 1955. The first version was published in 1961, and the Enzyme Commission was dissolved at that time, though its name lives on in the term EC Number. The current sixth edition, published by the International Union of Biochemistry and Molecular Biology in 1992 as the last version published as a printed book, contains 3196 different enzymes. Supplements 1-4 were published 1993-1999. Subsequent supplements have been published electronically, at the website of the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology.[5] In August 2018, the IUBMB modified the system by adding the top-level EC 7 category containing translocases.[9]
See also[edit]
- List of EC numbers
- List of enzymes
- TC number (classification of membrane transport proteins)
References[edit]
- ^ Webb, E. C. (1992). Enzyme nomenclature 1992: recommendations of the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology on the nomenclature and classification of enzymes. Academic Press. ISBN 978-0-12-227164-9.
- ^ a b «ENZYME (Enzyme nomenclature database)». ExPASy. Archived from the original on 21 March 2019. Retrieved 24 April 2019.
- ^ Omelchenko MV, Galperin MY, Wolf YI, Koonin EV (2010). «Non-homologous isofunctional enzymes: a systematic analysis of alternative solutions in enzyme evolution». Biology Direct. 5 (1): 31. doi:10.1186/1745-6150-5-31. PMC 2876114. PMID 20433725.
- ^ Apweiler R, Bairoch A, Wu CH, Barker WC, Boeckmann B, Ferro S, Gasteiger E, Huang H, Lopez R, Magrane M, Martin MJ, Natale DA, O’Donovan C, Redaschi N, Yeh LS (Jan 2004). «UniProt: the Universal Protein knowledgebase». Nucleic Acids Research. 32 (Database issue): D115–9. doi:10.1093/nar/gkh131. PMC 308865. PMID 14681372.
- ^ a b Moss GP. «Recommendations of the Nomenclature Committee». International Union of Biochemistry and Molecular Biology on the Nomenclature and Classification of Enzymes by the Reactions they Catalyse. Archived from the original on 2018-09-10. Retrieved 2006-03-14.
- ^ Rahman SA, Cuesta SM, Furnham N, Holliday GL, Thornton JM (Feb 2014). «EC-BLAST: a tool to automatically search and compare enzyme reactions». Nature Methods. 11 (2): 171–174. doi:10.1038/nmeth.2803. PMC 4122987. PMID 24412978.
- ^ Hoffman-Ostenhof, O (1953). «Suggestions for a more rational classification and nomenclature of enzymes». Advances in Enzymology and Related Areas of Molecular Biology. Advances in Enzymology and Related Subjects of Biochemistry. Vol. 14. pp. 219–260. doi:10.1002/9780470122594.ch7. ISBN 9780470122594. PMID 13057718.
- ^ Dixon, M; Webb, E.C. (1958). Enzymes. London: Longmans Green. pp. 183–227.
- ^ Tipton, Keith (August 2018). «Enzyme Nomenclature News: Translocases (EC 7): A new EC Class». ExplorEnz: the primary source of the IUBMB enzyme list. Archived from the original on 10 September 2018. Retrieved 3 November 2018.
External links[edit]
![]()
- Enzyme Nomenclature, authoritative website by the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology, maintained by G.P. Moss
- Enzyme nomenclature database — by ExPASy
- List of all EC numbers — by BRENDA
- Browse PDB structures by EC number
- Browse SCOP domains by EC number — by dcGO
- Compare EC numbers using EC-Blast Archived 2019-05-30 at the Wayback Machine
The Enzyme Commission number (EC number) is a numerical classification scheme for enzymes, based on the chemical reactions they catalyze.[1] As a system of enzyme nomenclature, every EC number is associated with a recommended name for the corresponding enzyme-catalyzed reaction.
EC numbers do not specify enzymes but enzyme-catalyzed reactions. If different enzymes (for instance from different organisms) catalyze the same reaction, then they receive the same EC number.[2] Furthermore, through convergent evolution, completely different protein folds can catalyze an identical reaction (these are sometimes called non-homologous isofunctional enzymes)[3] and therefore would be assigned the same EC number. By contrast, UniProt identifiers uniquely specify a protein by its amino acid sequence.[4]
Format of number[edit]
Every enzyme code consists of the letters «EC» followed by four numbers separated by periods. Those numbers represent a progressively finer classification of the enzyme. Preliminary EC numbers exist and have an ‘n’ as part of the fourth (serial) digit (e.g. EC 3.5.1.n3).[2]
For example, the tripeptide aminopeptidases have the code «EC 3.4.11.4», whose components indicate the following groups of enzymes:
- EC 3 enzymes are hydrolases enzymes (enzymes that use water to break up some other molecule)
- EC 3.4 are hydrolases that act on peptide bonds
- EC 3.4.11 are those hydrolases that cleave off the amino-terminal amino acid from a polypeptide
- EC 3.4.11.4 are those that cleave off the amino-terminal end from a tripeptide
Top level codes[edit]
| Class | Reaction catalyzed | Typical reaction | Enzyme example(s) with trivial name |
|---|---|---|---|
| EC 1 Oxidoreductases | Oxidation/reduction reactions; transfer of H and O atoms or electrons from one substance to another | AH + B → A + BH (reduced) A + O → AO (oxidized) | Dehydrogenase, oxidase |
| EC 2 Transferases | Transfer of a functional group from one substance to another. The group may be methyl-, acyl-, amino- or phosphate group | AB + C → A + BC | Transaminase, kinase |
| EC 3 Hydrolases | Formation of two products from a substrate by hydrolysis | AB + H2O → AOH + BH | Lipase, amylase, peptidase, phosphatase |
| EC 4 Lyases | Non-hydrolytic addition or removal of groups from substrates. C-C, C-N, C-O or C-S bonds may be cleaved | RCOCOOH → RCOH + CO2 or [X-A+B-Y] → [A=B + X-Y] | Decarboxylase |
| EC 5 Isomerases | Intramolecule rearrangement, i.e. isomerization changes within a single molecule | ABC → BCA | Isomerase, mutase |
| EC 6 Ligases | Join together two molecules by synthesis of new C-O, C-S, C-N or C-C bonds with simultaneous breakdown of ATP | X + Y + ATP → XY + ADP + Pi | Synthetase |
| EC 7 Translocases | Catalyse the movement of ions or molecules across membranes or their separation within membranes | Transporter |
Reaction similarity[edit]
Similarity between enzymatic reactions can be calculated by using bond changes, reaction centres or substructure metrics (formerly EC-BLAST], now the EMBL-EBI Enzyme Portal).[6]
History[edit]
Before the development of the EC number system, enzymes were named in an arbitrary fashion, and names like old yellow enzyme and malic enzyme that give little or no clue as to what reaction was catalyzed were in common use. Most of these names have fallen into disuse, though a few, especially proteolyic enzymes with very low specificity, such as pepsin and papain, are still used, as rational classification on the basis of specificity has been very difficult.
By the 1950s the chaos was becoming intolerable, and after Hoffman-Ostenhof[7] and Dixon and Webb[8] had proposed somewhat similar schemes for classifying enzyme-catalyzed reactions, the International Congress of Biochemistry in Brussels set up the Commission on Enzymes under the chairmanship of Malcolm Dixon in 1955. The first version was published in 1961, and the Enzyme Commission was dissolved at that time, though its name lives on in the term EC Number. The current sixth edition, published by the International Union of Biochemistry and Molecular Biology in 1992 as the last version published as a printed book, contains 3196 different enzymes. Supplements 1-4 were published 1993-1999. Subsequent supplements have been published electronically, at the website of the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology.[5] In August 2018, the IUBMB modified the system by adding the top-level EC 7 category containing translocases.[9]
See also[edit]
- List of EC numbers
- List of enzymes
- TC number (classification of membrane transport proteins)
References[edit]
- ^ Webb, E. C. (1992). Enzyme nomenclature 1992: recommendations of the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology on the nomenclature and classification of enzymes. Academic Press. ISBN 978-0-12-227164-9.
- ^ a b «ENZYME (Enzyme nomenclature database)». ExPASy. Archived from the original on 21 March 2019. Retrieved 24 April 2019.
- ^ Omelchenko MV, Galperin MY, Wolf YI, Koonin EV (2010). «Non-homologous isofunctional enzymes: a systematic analysis of alternative solutions in enzyme evolution». Biology Direct. 5 (1): 31. doi:10.1186/1745-6150-5-31. PMC 2876114. PMID 20433725.
- ^ Apweiler R, Bairoch A, Wu CH, Barker WC, Boeckmann B, Ferro S, Gasteiger E, Huang H, Lopez R, Magrane M, Martin MJ, Natale DA, O’Donovan C, Redaschi N, Yeh LS (Jan 2004). «UniProt: the Universal Protein knowledgebase». Nucleic Acids Research. 32 (Database issue): D115–9. doi:10.1093/nar/gkh131. PMC 308865. PMID 14681372.
- ^ a b Moss GP. «Recommendations of the Nomenclature Committee». International Union of Biochemistry and Molecular Biology on the Nomenclature and Classification of Enzymes by the Reactions they Catalyse. Archived from the original on 2018-09-10. Retrieved 2006-03-14.
- ^ Rahman SA, Cuesta SM, Furnham N, Holliday GL, Thornton JM (Feb 2014). «EC-BLAST: a tool to automatically search and compare enzyme reactions». Nature Methods. 11 (2): 171–174. doi:10.1038/nmeth.2803. PMC 4122987. PMID 24412978.
- ^ Hoffman-Ostenhof, O (1953). «Suggestions for a more rational classification and nomenclature of enzymes». Advances in Enzymology and Related Areas of Molecular Biology. Advances in Enzymology and Related Subjects of Biochemistry. Vol. 14. pp. 219–260. doi:10.1002/9780470122594.ch7. ISBN 9780470122594. PMID 13057718.
- ^ Dixon, M; Webb, E.C. (1958). Enzymes. London: Longmans Green. pp. 183–227.
- ^ Tipton, Keith (August 2018). «Enzyme Nomenclature News: Translocases (EC 7): A new EC Class». ExplorEnz: the primary source of the IUBMB enzyme list. Archived from the original on 10 September 2018. Retrieved 3 November 2018.
External links[edit]
![]()
- Enzyme Nomenclature, authoritative website by the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology, maintained by G.P. Moss
- Enzyme nomenclature database — by ExPASy
- List of all EC numbers — by BRENDA
- Browse PDB structures by EC number
- Browse SCOP domains by EC number — by dcGO
- Compare EC numbers using EC-Blast Archived 2019-05-30 at the Wayback Machine
Международная классификация ферментов
- Международная классификация ферментов
-
Шифр КФ (Классификация ферментов) или код фермента — это классификационный номер фермента по международной иерархической классификации. Принятая система классифицирует ферменты по группам и индексирует индивидуальные ферменты, что важно для стандартизации исследований.
Классификация ферментов основана и периодически обновляется Комиссией по ферментам (англ. Enzyme commission, отсюда термин «EC number», принятый в англоязычной литературе) при Международном союзе биохимии и молекулярной биологии. Каждый шифр КФ ассоциирован также с рекомендованным названием соответствующего фермента. Классифицировано более 3500 ферментов.
Содержание
- 1 Принцип классификации
- 2 Формат шифра
- 2.1 Класс
- 2.2 Подкласс
- 2.3 Под-подкласс
- 2.4 Код четвертого уровня
- 3 Коды первого уровня
- 4 История
- 5 Ссылки
- 6 См.также
Принцип классификации
Классификация ферментов учитывает реакционную и субстратную специфичности ферментов, а не их белковую структуру. Шифр КФ определяет химическую реакцию, катализируемую ферментом. По этой причине аналогичные ферменты (иногда десятки) из различных организмов имеют один КФ, несмотря на структурные различия.
Иногда различные ферменты одного организма имеют один КФ. Например, панкреатическая липаза и печёночная липаза принадлежат обе к КФ 3.1.1.3 благодаря катализированию одной химической реакции (гидролиз эфирной связи в триглицериде), хотя первый фермент является ферментом пищеварения и работает в кишечнике, а второй относится к ферментам обмена липопротеинов в крови.
Существует база данных UniProt [1] , которая идентифицирует каждый белок по его первичной последовательности. Обе базы данных дополняют друг друга.
Формат шифра
Каждый классификационный номер содержит сокращение КФ и последовательность из четырёх чисел, разделённых точкой, и составляется по определенному принципу. Каждое последующее число представляет собой всё более и более уточняющую классификацию фермента. Так как база данных постоянно обновляется коды могут меняться и некоторые коды могут оставаться незаполненными.
Класс
- Код первого уровня (первое число, может быть от 1 до 6) указывает номер одного из шести главных классов ферментов (см. таблицу Коды первого уровня).
Подкласс
- Код второго уровня (второе число) означает подкласс, характеризующий основные виды субстратов, участвующих в данном типе химических реакций. Например, у трансфераз вторая цифра указывает на природу той группы, которая подвергается переносу, у гидролаз — на тип гидролизуемой связи и т. д.
Под-подкласс
- Код третьего уровня (третье число) определяет более частные подгруппы (под-подклассы), отличающиеся природой химических соединений доноров или акцепторов, участвующих в данной подгруппе реакций. У гидролаз, например, эта цифра уточняет тип гидролизуемой связи, а у лиаз — тип отщепляемой группы и т. д. Первые 3 числа шифра точно определяют тип фермента.
Код четвертого уровня
Наконец, все ферменты, относящиеся к данному под-подклассу, получают свой порядковый номер (четвёртое число в шифре).
- Например, глюкозоксидазе присвоен шифр КФ 1.1.3.4, что означает:
- КФ 1 — Оксидоредуктазы
- КФ 1.1 — Алкогольоксидоредуктазы
- КФ 1.1.3 — Оксидоредуктазы, окисляющие группу CH-OH и восстанавливающие кислород.
- КФ 1.1.3.4 — Оксидоредуктазы, окисляющие глюкозу в присутствие кислорода. Всего известно 3 глюкозоксидазы из разных организмов.
Коды первого уровня
Класс Катализируемая реакция Тип реакции Важнейшие подклассы КФ 1
ОксидоредуктазыОкислительно-восстановительные реакции. Перенос атомов H и O или электронов от одного субстрата на другой AH + B → A + BH (восстановленный)
A + O → AO (окисленный)дегидрогеназа, оксидаза, пероксидаза, редуктаза, монооксидаза, диоксигеназа КФ 2
ТрансферазыПеренос функциональной группы от одного субстрата на другой. Это может быть метильная, ацильная, фосфатная группа или аминогруппа. AB + C → A + BC аминотрансфераза, фосфотрансфераза, C1-трансфераза, гликозилтрансфераза КФ 3
ГидролазыОбразование двух продуктов из одного субстрата в результате гидролиза. AB + H2O → AOH + BH эстераза, гликозил-гидролаза, пептидаза, амидаза КФ 4
Лиазы (синтазы)Негидролитическое добавление или удаление группы к или от субстрата. Образование C-C, C-N, C-O или C-S связи. RCOCOOH → RCOH + CO2 C-O-лиаза, C-S-лиаза, C-N-лиаза, C-C-лиаза КФ 5
ИзомеразыВнутримолекулярная перестановка, то есть изомеризация молекулы субстрата. AB → BA эпимераза, цис-транс-изомераза, внутримолекулярная оксидоредуктаза и др. КФ 6
Лигазы (синтетазы)Соединение двух молекул в результате синтеза новой C-O, C-S, C-N или C-C связи, сопряжённое с одновременным гидролизом АТФ. X + Y+ ATP → XY + ADP + Pi C-O-лигаза, C-S-лигаза, C-N-лигаза, C-C-лигаза История
Схема номенклатуры ферментов была впервые разработана в 1955 году, когда Международный конгресс биохимии в Брюсселе учредил Комиссию по ферментам (Enzyme Commission). Первая версия номенклатуры появилась в 1961 году и включала около 900 ферментов, в версии 1978 года было более 2000 ферментов. Версия 1995 года содержит более 3500 ферментов.
Ссылки
- Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (NC-IUBMB), Официальный сайт
- Список ферментов
- ↑ ENZYME (Enzyme nomenclature database). ExPASy. Проверено 14 марта 2006.
См.также
- Ферменты
- Список ферментов
- Кластер дифференцировки
Шаблон:Ферменты
Wikimedia Foundation.
2010.
Полезное
Смотреть что такое «Международная классификация ферментов» в других словарях:
-
классификация ферментов — КФ Международная четырехзначная номенклатура ферментов: 1 й знак класс ферментов (1 оксиредуктазы; 2 трансферазы; 3 гидролазы; 4 лиазы; 5 изомеразы; 6 лигазы); 2 й и 3 й знаки подклассы и подподклассы ферментов; 4 й знак порядковый номер в… … Справочник технического переводчика
-
классификация ферментов — Enzyme Classification (EC) классификация ферментов (КФ). Международная четырехзначная номенклатура ферментов: 1 й знак класс ферментов (1 оксиредуктазы; 2 трансферазы; 3 гидролазы; 4 лиазы; 5 изомеразы; 6 лигазы); 2 й и 3 й знаки подклассы и… … Молекулярная биология и генетика. Толковый словарь.
-
Приложение. Список сокращений — a. arteria (ед. число) aa. arteriae (мн. число) ant. anterior b. bursa (ед. число) Bac. Bacillus Bact. Bacterium bb. bursae (мн. число) Ber определитель бактерий Берджи (Bergey’s manual of determinative bacteriology, 8 ed., 1974) BNA Базельская… … Медицинская энциклопедия
-
МКБ-10: Класс III — Международная классификация болезней 10 го пересмотра (МКБ 10) Класс I Некоторые инфекционные и паразитарные болезни Класс II Новообразования Класс III Болезни крови, кроветворных ор … Википедия
-
МКБ-10: Класс III. Болезни крови, кроветворных органов и отдельные нарушения, вовлекающие иммунный механизм — Международная классификация болезней 10 го пересмотра (МКБ 10) Класс I Некоторые инфекционные и паразитарные болезни Класс II Новообразования Класс III Болезни крови, кроветворных органов и отдельные нарушения, вовлекающие иммунный механизм Класс … Википедия
-
МКБ-10: Класс XX — Международная классификация болезней 10 го пересмотра (МКБ 10) Класс I Некоторые инфекционные и паразитарные болезни Класс II Новообразования Класс III Болезни крови, кроветворных органов и отдельные нарушения, вовлекающие иммунный ме … Википедия
-
МКБ-10: Код Y — Международная классификация болезней 10 го пересмотра (МКБ 10) Класс I Некоторые инфекционные и паразитарные болезни Класс II Новообразования Класс III Болезни крови, кроветворных органов и отдельные нарушения, вовлекающие иммунный механизм Класс … Википедия
-
Хронический панкреатит — МКБ 10 K86.086.0 K86.186.1 МКБ 9 577.1 … Википедия
-
Инфекцио́нные боле́зни — (позднелат. infectio заражение) группа болезней, которые вызываются специфическими возбудителями, характеризуются заразительностью, циклическим течением и формированием постинфекционного иммунитета. Термин «инфекционные болезни» был введен… … Медицинская энциклопедия
-
МКБ-10: Класс IV — Список классов Международной классификации болезней 10 го пересмотра Класс I. Некоторые инфекционные и паразитарные болезни Класс II. Новообразования Класс III. Болезни крови, кроветворных органов и отдельные нарушения, вовлекающие иммунный… … Википедия
-
Классификация и номенклатура ферментов
Согласно
классификации, разработанной Международной
комиссией по ферментам и принятой в
1961 году, все ферменты разделяют на шесть
классов в зависимости от характера
катализируемых реакций:
-
Оксидоредуктазы.
Катализируют окислительно-восстановительные
реакции. -
Трансферазы.
Катализируют реакции переноса группировок
от одного соединения на другое. -
Гидролизы.
Ускоряют гидролитическое расщепление
веществ. -
Лиазы.
Катализируют реакции негидролитического
расщепления с образованием двойных
связей или реакции присоединения групп
к двойным связям. -
Изомеразы.
Катализируют реакции изомеризации
соединений. -
Лигазы
(синтетазы). Ускоряют реакции синтеза
с использованием энергии макроэргических
соединений.
Классы
делятся на подклассы,
последние — на подподклассы
или группы, внутри которых ферментам
присвоен порядковый номер. Каждый
фермент имеет свой индивидуальный
четырехзначный шифр, первая цифра
которого обозначает класс, вторая —
подкласс, третья — подподкласс, четвертая
— порядковый номер фермента.
Ферменты
могут иметь систематические (рациональные)
и рабочие (тривиальные) названия.
Систематические названия, предложенные
Международной комиссией, включают
названия субстратов (их указывают через
двоеточие) и характер катализируемых
реакций. Если фермент катализирует
реакцию, при которой одновременно идут
два превращения, то в наименовании
фермента второе превращение дается в
скобках.
-
Характеристика классов ферментов
Оксидоредуктазы
[1]
Оксидоредуктазы
— ферменты катализирующие
окислительно-восстановительные реакции.
При их участии протекают такие важнейшие
физиологические процессы как дыхание
и фотосинтез, происходят превращения
жирных кислот, фенольных соединений,
витаминов и других веществ.
Окислительно-восстановительные
реакции в живых организмах осуществляются
за счет переноса протонов, электронов
или присоединения кислорода.
Оксидоредуктазы, катализирующие перенос
водорода, имеют тривиальное название
дегидрогеназы.
Систематические названия ферментов
этого класса составляют по схеме: донор
водорода : акцептор
водорода — оксидоредуктаза.
В тех случаях, когда донор водорода
точно не установлен, используют термин
редуктаза.
Реакции прямого присоединения кислорода
к субстрату катализируют ферменты
оксигеназы.
Все ферментативные ОВР обратимы.
Класс
оксидоредуктазы подразделяется на
подклассы в зависимости от того, какие
группировки подвергаются окислению. В
основу деленния на подподклассы положен
химизм акцептора водорода (таблица 1).
Таблица
1.
Ферменты
класса Оксидоредуктазы
| Подкласс | Подполклас | Название |
| 1.1. | 1.1.1. | алкогольдегидрогеназа малат изоцитрат лактат глюкозо-6-фосфат глицерин-3-фосфат |
| 1.1.3. | глюкоза | |
| 1.2. | 1.2.1. | глицеральдигид-фосфат |
| 1.2.4. | пируват -кетоглутарат | |
| 1.3. | 1.3.1. | Еноил-АПБ-редуктаза |
| 1.3.99. | Сукцинатдегидрогеназа | |
| 1.4. | 1.4.1. | Глутамат |
| 1.6. (действующие | (с | Дегидрогеназы |
| 1.9. | 1.9.3. | Цитохром |
| 1.10. (действующие | 1.10.2. Сu-протеиды | Убихинон-цитохром Ортодифенолоксидаза Парадифенолоксидаза Аскорбинатоксидаза |
| 1.11. Действущие | Fe-протеиды | Каталаза, Пероксидаза |
| 1.13. Действующие | 1.3.11. | Триптофан липоксигеназа |
| 1.14. | 1.14.16. | Фенилаланин |
| 1.17. | 1.17.4. | Рибонуклеозид-дифосфат |
| 1.18. | Нитрогеназа |
Дегидрогеназы
— двухкомпонентные ферменты, имеющие
пиридиновые (НАД+
и
НАДФ+)
и флавиновые (ФАД
и
ФМН)
коферменты. НАД+
(никотинамиддинуклеотид)
имеет в составе одного нуклеотида
пиридиновое производное — амид никотиновой
кислоты (витамин РР) и рибозу, в состав
другого входит азотистое основание
аденин и рибоза (или фосфорный эфир
рибозы). Оба нуклеотида соединены через
остатки фосфорной кислоты. В состав
НАД+
и НА.ДФ+
находятся во всех типах клеток, причем
НАД содержится обычно в значительно
больших количествах по сравнению с
НАДФ+.
НАДФ+
входит не два, а три остатка фосфорной
кислоты (рисунок 1).
|
| Амид | |
| Остатки | Рибоза | |
|
Аденин Рибоза | ||
| Рисунок |





Каталитические
свойства НАД+—
и НАДФ+— содержащих
дегидрогеназ определяются пиридиновым
кольцом, к одному из углеродных атомов
которого при восстановлении присоединяются
один протон и один электрон (атом
водорода). Второй электрон присоединяется
к положительно заряженному атому азота.
Остающийся протон переходит в окружающую
среду и используется затем в последующих
реакциях. Восстановленную форму НАД+
и НАДФ+
обозначают соответственно НАД·Н
+ Н+
и НАДФ·Н
+ Н+.
Коферменты
ФАД
(FAD)
— флавинадениндинуклеотид и ФМН
(FMN)
— флавинмононуклеотид — являются
производными витамина
В2
— рибофлавина. Флавиновые ферменты
связывают водород, присоединяя два
атома водорода к двум атомам азота в
изоаллоксазиновой группировке с
соответствующей перегруппировкой
двойных связей (рисунок 2, 3).
|
|
|
| Витамин | Остаток |
| Рисунок 2. Флавинадениндинуклеотид | Рисунок |





Открыто
более 80 флавиновых ферментов. Они
катализируют окисление полуацеталей
в лактоны, спиртов в альдегиды, аминов
в имины, насыщенных ЖК в ненасыщенные
ЖК, НАДН
в НАД+.
В состав флавиновых ферментов могут
входить металлы, образуя гем
(альдигидоксилаза,
ксантиноксидаза)
или железоцентры.
По
характеру действия дегидрогеназы
делятся на анаэробные и аэробные
(оксидазы). Аэробные передают отщепляемый
от окисляемого субстрата водород
непосредственно на кислород воздуха,
а анаэробные — на промежуточные переносчики
(обычно аэробные дегидрогеназы).
Пиридиновые дегидрогеназы — ферменты
анаэробные, среди флавиновых встречаются
как анаэробные так и аэробные (оксидазы).
Ниже
приведены примеры ферментов, участвующих
в некоторых окислительно-восстановительных
превращениях, лежащих в основе фотосинтеза,
дыхания, брожения и др.
Глицеральдигид-3-фосфат
дегидрогеназа (1.2.1.12)
катализирует одну из важнейших реакций
гликолиза (анаэробного этапа дыхания)
-окисление глицеральдигид-3-фосфата до
1,3-дифосфоглицериновой кислоты по схеме:

Фермент
является олигомером — состоит из 4-х
субъединиц, каждая из которых содержит
НАД+
и SH-группу
(радикал цистеина), входящую в состав
активного центра. Реакция происходит
через промежуточное образование тиоэфира
между окисляемой альдегидной группой
и SH-группой.
Малатдегидрогеназа
(1.1.1.37) в цикле Кребса катализирует
превращение малата (яблочной кислоты)
в оксалацетат (щавелевоуксусную кислоту):

Изоцитратдегидрогеназа
(1.1.1.42) — один из ферментов цикла Кребса,
катализирующий превращение изоцитрата
(изолимонной кислоты) в щавелевоянтарную
кислоту:
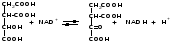
*Глицерол-3-фосфат-дегидрогепаза
(1.1.1.8) катализирует превращение
глицерол-З-фосфата в фосфодиоксиацетон.:

В
обратной реакции образуется
глицерол-З-фосфат, используемый в
биосинтезе фосфоглицеридов и жиров.
*Алкогольдегидрогеназа
(1.1.1.2) катализирует превращение этилового
спирта в уксусный альдегид (способствует
окислению пива, вина и других
спиртосодержащих напитков):
СН3СН2ОН
+ NADF+
СН3СОН
+ NADF
Н + Н+
Сукцинатдегидрогеназа
(1.3.99.1),
является флавиновой анаэробной
дегидрогеназой, катализирующей в цикле
Кребса окисление сукцината (янтарной
кислоты) до фумарата (фумаровой кислоты):
СООН-СН2-СН2-СООН
+ FAD
СООН-СН=СН-СООН + FAD.Н2
В
цепи окислительно-восстановительных
реакций, например, при окислительном
фосфорилировании, восстановленные
формы флавиновых дегидрогеназ могут
передавать водород не только кислороду,
но также полифенолоксидазный и цитохромной
системам.
* Полифенолоксидаза
(1.14.18.1)
является Сu-протеидом,
окисляющим моно-, ди-, и трифенолы в
соответствующие хиноны. содержится в
грибах и высших растениях. Потемнение
поверхности разрезанного яблока или
клубня картофеля, плодов и овощей при
сушке, при скручивании и завядании
чайного листа объясняется окислением
полифенолов и дубильных веществ
полифенолоксидазой.

Полифенолоксидазой
можно назвать фермент * тирозиназу,
окисляющую аминокислоту тирозин с
образованием темно-окрашенных соединений
— меланинов (в грибах, ржаной муке).
Потемнение макарон при сушке также
объясняется высокой активностью
тирозиназы в некоторых партиях пшеничной
муки.
Цитохромная
система
включает более двадцати цитохромов (a,
b,
c
и др.) и цитохромоксидазу (a3).
Цитохромы
состоят из железопорферина (гема),
связанного с белком (рис. 3). Все они
участвуют в окислительно-восстановительных
реакциях (в окислительном и фотосинтетическом
фосфорилировании) и служат переносчиками
электронов.
Fe3+
+ e—
Fe2+
Цитохромоксидаза
(1.9.3.1) активирует кислород путем
присоединения к нему электронов. Является
сложным ферментом, содержащим Сu.
* Пероксидаза
(1.11.1.7.) — железосодержащий фермент,
осуществляющий окисление органических
соединений перекисью водорода. При этом
образуются комплексные соединения,
перекись активируется и приобретает
способность действовать как акцептор
водорода. Пероксидаза окисляет полифенолы
и некоторые ароматические амины. В
качестве перекиси могут выступать
соединения с двойными связями при их
окислении: терпены, каротиноиды,
ненасыщенные жирные кислоты.
Рисунок
3. Простетическая группа цитохрома
с.
* Каталаза
(1.11.1.6.) расщепляет перекись водорода на
воду и кислород. Каталаза содержит Fe
в составе гематина, её ингибируют HCN,
H2S,
фториды.
* Аскорбинатоксидаза
(1.10.3.3)
— Сu-протеид,
содержащий до 0,24 %
меди, окисляет аскорбиновую кислоту
(витамин С) до дегидроаскорбиновой
кислоты. Особенно активна аскорбинатоксидаза
в тыкве, капусте, кабачках.

* Альдегидоксидаза
( ) относится к флавиновым ферментам,
она окисляет уксусный альдегид до
уксусной кислоты:
СН3-СОН
+ Н2О
+ О2
СН3-СООН
+ Н2О2
Лактатоксидаза
(
) —
флавиновый
фермент, содержащийся
в … и окисляющий
лактат (молочную кислоту) в ацетат
(уксусную кислоту):
СН3
СНОН СООН + О2
СН3-СООН
+ СО2
+ Н2О
* Глюкозооксидаза
(1.1.3.4)
содержится в различных плесневых
грибах. Она получена в кристаллическом
виде, содержит две молекулы ФАД и 15 %
углеводов. Фермент легко окисляет
глюкозу до глюконовой кислоты.
* Липоксигеназа
(липоксидаза) (1.13.11.12) окисляет некоторые
ненасыщенные жирные кислоты и образуемые
ими сложные эфиры. Этот фермент хорошо
окисляет линолевую и линоленовую кислоты
с образованием гидроперекисей, которые
потом сами могут окислять другие жирные
кислоты, а также каротиноиды и витамин
А, аминокислоты, хлорофилл, витамин С.
Эти процессы происходят во время сушки
и хранения различных растительных
продуктов. Прогоркание муки и круп
связано с действием липоксигеназы.
* Нитратредуктаза
(1.6.6.1) и нитритредуктаза
(1.6.6.4)
играют очень важную роль в азотном
обмене растений. Первый восстанавливает
нитраты до нитритов, а второй — нитриты
до аммиака. Донором протонов и электронов
в этих реакциях является НАДН
(или НАДФН):
HNO3
+ NADH
+ H+
HNO2
+ H2O
+ NAD+
HNO2
+ 3 NADH
+ 3 H+
NH3
+ 2 H2O
+ NAD+
Нитратредуктаза
состоит из двух частей, диафоразной,
содержащей ФАД, и терминальной, содержащей
молибден. Нитритредуктаза
являются
флавинсодержащим железосерным белком.
Таким
образом, железо и медь, входящие в состав
многих оксидаз, при наличии их в среде
(используемая для переработки или
хранения посуда), способствуют быстрому
окислению, потемнению и порчи продукции
растениеводства. Однако снижение
активности окислительно-восстановительных
реакций в растениях из-за недостатка
этих микроэлементов, снижает их
устойчивость к болезням, урожайность
и качество продукции. Недостаток в почве
таких микроэлементов как молибдена,
магния, марганца, железа и меди, являющихся
активаторами ферментов
нитрат-восстанавливливающей системы
растений, приводит к снижению содержания
белков и накоплению токсичных для
человека и животных нитратов и нитритов.
Трансферазы [2]
Трансферазы —
ферменты, катализирующие реакции
переноса отдельных групп или молекул.
Систематическое
название ферментов этого класса
образуется следующим образом: донор
акцептор — транспортируемая группа —
трансфераза. Часто используют н цифровую
приставку для указания положения, к
которому присоединяется переносимая
группа.
Выделено 8 подклассов
— по химизму переносимых групп.
2.1. Переносят
одноуглеродные
остатки: метильные, оксиметильные,
формильные, карбоксильные и др. Пример:
метилтрансферазы,
катализирующие перенос метальных групп,
принимают участие в синтезе холина
(лецетины), пуринов, пиримидинов, НК.
Коферментами являются фолиевая
к-та и вит.
В12.
2.2. Переносят
альдегидные
и котонные
остатки. Сюда относятся ферменты:
транскетолаза
(переносит 2-углеродный фрагмент) н
трансальдолаза
(переносит 3-углеродный фрагмент). Эти
ферменты участвуют в пентозном цикле
дыхания, в фотосинтетическом цикле
Кальвина.
2.3. Ацилтрансферазы
— ферменты, переносящие остатки уксусной
к-ты СН3СО-,
а также остатки жирных кислот. Это
двухкомпонентные ферменты, в состав
которых входит кофермент А. При переносе
остатков уксусной кислоты в реакции
участвует ацетилкофермент
А
(СН3СОS-КоА),
а при переносе остатков жирных кислот
— ацилкофермент
А (R-СОS-КоА).
С их участием идет синтез лимонной
к-ты, жирных
кислот, каучука
и др. в-в. ацетилглюкозамина, ацетилхолина.
2.4. Гликозилтрансферазы
— переносчики остатков моносахаридов.
Эти ферменты участвуют в синтезе
полисахаридов и имеют в качестве
небелковой части нуклеозиддифосфатсахара
(НДФС). Показать
рисунок !
К числу
гликозилтрансфераз относят ферменты,
которые были известны под названием
фосфоризаз,
широко распостраненных у растений,
животных и микроорганизмах. Представителем
является крахмальная фосфорилаза —
-глюканфосфорилаза
(2.4.1.1), катализирующая превращение
крахмала в гл.-1-фосфат, перенос глюкозы
на фосфат:
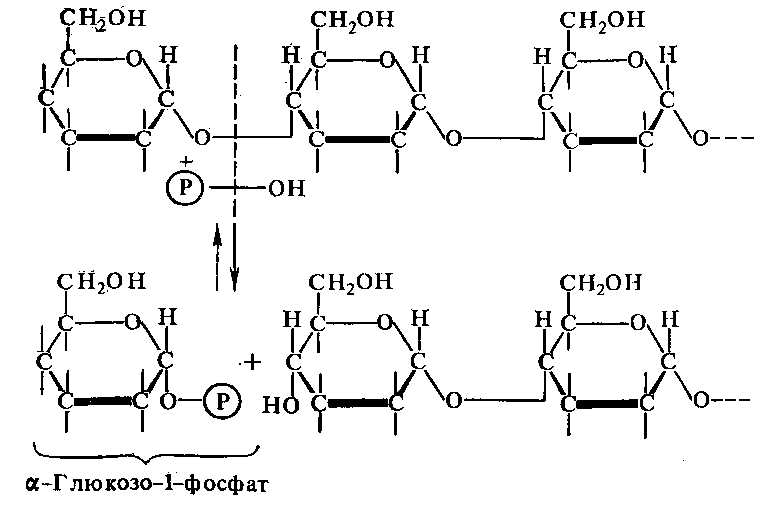
Фосфоролитическое
расщепление нуклеозидов также идет при
участии этих ферментов.
2.6. Переносят группы,
содержащие азот.
К этому подклассу относятся важнейшие
ферменты азотного обмена — аминотрансферазы.
Аминотрансферазы катализируют
межмолекулярный перенос аминогруппы
между аминокислотами и кетокислотами
в процессе реакции переаминирования,
идущими по схеме:
R1 R2 R1 R2
CHNH2
+ CO = CO + CHNH2
COOH COOH COOH COOH
Аминотрансферазы-
двухкомпонентные ферменты, активной
группой которых является фосфорилированное
производное
витамина
В6
(пиридоксина).

2.7. Переносят группы,
содержащие фосфор
(фосфатрансферазы или киназы).
Эти ферменты переносят фосфатный остаток
от АТФ на различные акцепторы, пирофосфатный
остаток пирофосфотрансферазы), а также
более сложные группировки, содержащие
фосфатный остаток (нуклеотидилтрансферазы
и др.).
Фермент гексокиназа
(2.7.1.1) ускоряет
фосфорилирование гексоз (глюкозы и
фруктозы).
Фермент
фосфофруктокиназа
(2.7.1.11)
катализирует фосфорилирование
фруктозо-6-фосфата во фруктозо-1,6-дифосфат
(смотри уч.: Плешков, 1987; стр.68)
Фермент фосфоглюкомутаза
(2.7.5.1) катализирует взаимное обратимое
превращение глюкозо-1-фосфата и
глюкозо-6-фосфата

2.8. Переносят группы,
содержащие серу. В этот подкласс входят,
в частности, КоА-трансферазы,
переносящие остатки кофермента А.
Гидролизы [3]
Ферменты этого
класса катализируют гидролиз, а иногда
и синтез сложных соединений с участием
воды:
RR1
+ HOH
= ROH
+ R1H
Систематическое
название ферментов класса гидролаз
составляется из названия субстрата и
через тире слова гидролиза. Отщепляемые
группы могут быть указаны после названия
субстрата, например аденозин —
аминогидролаза. Класс гидролаз делится
на одиннадцать подклассов.
3.1. Гидролизуют
сложноэфирные связи
(эстеразы, фосфатазы, нуклеазы).
3.1.1.
Липазы
— катализируют
расщепление
жиров —
триацилглицеринов по схеме:

где R1,
R2
и R3
— остатки жирных кислот.
В растениях липазы
широко распространены: обмен жиров в
клетках в значительной степени связан
с активностью этих ферментов. У каждого
вида растений есть свои собственные
липазы, значительно различающиеся по
свойствам (растворимость, оптимум рН и
т. д.), однако в отличие от многих других
ферментов специфичность липаз очень
низка и любая липаза может расщеплять
всякий жир. Действие липазы имеет большое
значение при хранении муки и круп,
особенно содержащих большое ко-во жира
(пшено). При повышенной влажности этих
продуктов и повышенной температуре
хранения липаза быстро расшепляет
глицериды с образованием свободных
жирных кислот, что приводит к повышению
кислотности и его быстрому прогорканию.
К эстеразам относят
также пектинэстеразу или пектазу
(3.1.1.11) — осуществляющих гидролитическое
расщепление пектиновых вв. до галактуроновой
к-ты и метилового спирта.
3.1.2.Г и д р о л а з
ы т и о л о в ы х э ф и р о в.
Например, ацетил-
КоА-гидролаза
(3.4.2.1), катализирующая расщепление
ацетилкофермента А:
СН3СОS—KoA
+ H2O
СН3СOОH
+ HS—KoA
3.1.3. Гидролазы
фосфомоноэфиров
катализируют отщепление остатка
фосфорной кислоты от соединений типа
R
— 0- P. К таким соединениям относят,
например, фосфорные эфиры сахаров —
глюкоза-б-фосфат, глюкоза-1-фосфат,
фруктоза- 1 ,6 -дифосфат и др. Соответствующие
ферменты носят названия глюкозо-6-фосфатаза
(3.1.3.9), глюкоза-1-фосфатаза
(3.1.3.10), гексозодифосфатаза,
(3.1.3.11) н т. д. Схемы реакций, катализируемые
этими ферментами, можно представить
следующим образом:
3.1.4. Гидролазы
фосфодиэфиров.
К этой группе ферментов относятся
дезоксирибонуклеаза,
(3.1.4.5) и рибонуклеаза
(3.1.4.9), которые катализируют расщепление
соответственно ДНК и РНК.
3.2. Гидролизуют
гликозильные связи.
Среди этих ферментов в растениях наиболее
распространены гидролизы гликозидов,
катализирующие гидролитическое
расщепление
углеводов и гликозидов.
К этим ферментам относятся —
и -амилазы
(3.2.1.1 и 3.2.1.2), ускоряющие расщепление
крахмала (-амилаза
содержится
в слюне, пищеварительном соке выделяемом
поджелудочной железой, проросшем зерне
пшеницы, ржи, ячменя), целлюлаза
(3.2.1.4), катализирующая расщепление
целлюлозы, инулаза,
(3.2.1.7),
расщепляющая полисахарид инулин,
полигалактуроназа,
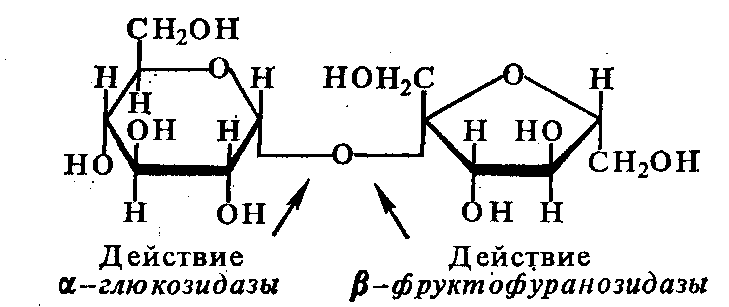
-глюкозидаза
(3.2.1.20), расщепляющая -гликозидную
связь в дисахаридах и гликозидах
(тождественнен мальтазе,
превращающей дисахарид мальтозу в две
молекулы глюкозы), -глюкозидаза
3.2.1.21),
расщепляющая ди- и полисахариды и
гликозиды (целобиозу, амигдалин, арбутин
и др.),
-фруктофуранозидаза
(сахараза,
инвертаза)
(3.2.1.26), катализирующая распад сахарозы,
-галактозидаза
(3.2.1.23), участвующая в расщеплении
дисахарида лактозы на молекулы галактозы
и глюкозы, хитиназа.
Схему действия гидролаз гликозидов
можно представить на примере гидролиза
сахарозы:
сахароза + H2O
= глюкоза + фруктоза.

3.4. Гидролизуют
пептидные связи
(пептид-гидролазы).
Систематической
номенклатуры для ферментов данного
подкласса нет, так как специфичность
этих ферментов перекрывается. В этом
подклассе различают ряд подподклассов
ферментов, в частности отщепляющие
N-концевые
аминокислоты (аминопептидазы),
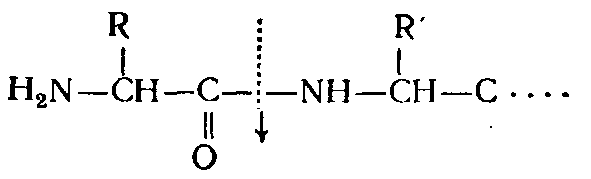 С-концевые аминокислоты (карбоксипептидазы),
С-концевые аминокислоты (карбоксипептидазы),
 гидролизующие дипептиды (дипептидазы),
гидролизующие дипептиды (дипептидазы),
несколько подподклассов пептид-гидролаз,
расщепляющих белки (пептидил-пептидгидролазы,
протеиназы
или протеолитические
ферменты ).
Пептидазы
(3.4.) или протеолитические ферменты,
катализируют гидролитическое расщепление
пептидных — С-N-
связей в белках или пептидах по схеме:
RCO—NHR1
+ H2O
= RCOOH
+ R1NH2
где R
и R1
— остатки аминокислот или пептидов.
Субстратами пептидаз
являются белки и пептиды, которые
расщепляются на пентиды меньшей
молекулярной массы или до свободных
аминокислот.
Известно несколько
десятков ферментов этого типа, из которых
наиболее хорошо изучены ферменты
желудочного сока — пепсин
( 3 4.4.1) , трипсин
( 3.4.4.4) и химотрипсин
(3.4. 4 .5 ) , а
также растительный фермент папаин
(3.4.4.10). Все они представляют собой
однакомпонентные ферменты; получены в
кристаллическом состоянии и достаточно
хорошо изучены. Протеиназы являются
эндопептидазами и гидролизуют белки
до пептидов разной величины. В зависимости
от строения активного центра выделяют
следующие подклассы протеиназ.
3.4.21. Сериновые
протеиназы — содержат в активном центре
серин, необходимый для катализа. Сюда
относятся химотрипсин,
трипсин —
протеиназы желудочно-кишечного тракта,
тромбин, плазмин — протеиназы крови.
3.4.22. Тиоловые
протеиназы — содержат в активном центре
цистеин. Это, в основном, растительные
протеиназы, например папаин
из млечного сока дынного дерева. Некоторые
тканевые протеиназы (катепсин В и др.).
3.4.23. Кислые протеиназы
— содержат в активном центре две
карбоксильные группы, оптимум рН их
действия от 1 до 5. К этому подклассу
принадлежат пепсин,
реннин
(химозин), катепсин D,
пеницилопепсин.
3.4.24. Металлопротеиназы.
3.4.99. Протеиназы с
неизвестным механизмом катализа.
3.5. Гидролизуют
С-N-связи,
отличные от пептидных.
В частности, сюда относятся ферменты,
расщепляющие амидные связи — дезамидазы.
Дезамидазы
(3.5.1 и 3.5.3) катализируют гидролитическое
дезаминирование амидов. Реакция идет
по схеме:
![]()
Эти ферменты играют
важную роль в азотном обмене.
Аспарагиназа
(3.5.1.1) и глутаминаза
(3.5.1.2) — ферменты, которые катализируют
гидролитическое расщепление соответственно
аспарагина и глутамина:

К этой группе
ферментов относится уреаза
(3.5.1.5), которая катализирует гидролитическое
расщепление мочевины:
![]()
Уреаза
(3.5.1.5) содержится во многих растениях,
особенно ее много семенах сои. Она очень
специфична, действует только на мочевину.
Уреаза — первый фермент, который был
получен в кристаллическом состоянии.
Дезаминазы
(3.5.3. и 3.5.4.) — ферменты, катализирующие
гидролитическое дезаминирование
азотистых оснований и нуклеотидов,
входящих в состав нуклеиновых кислот,
а также дезаминирование некоторых
других соединений. Пой действием
дезаминаз реакции идут по следующей
схеме:
![]()
Аргиназа
(3.5.3.1) катализирует гидролитическое
расщепление Д-аргинина на орнитин и
мочевину. Оптимальная тзона для действия
аргиназы при рН 10. Активируется солями
марганца (возможно содержит).
3.6. Гидролизуют
кислотно-ангидридные связи.
В этот подкласс входят неорганическая
полифосфатазы, катализирующие отщепление
фосфорной кислоты от полифосфатов.
Среди этих ферментов важную роль играют
пирофосфатаза
(3.6.1.1) действующая но схеме:
H4P2O7
+ H2O
2H3PO4
аденозинтрифосфатаза
или АТФаза
(3.6.1.3), катализирующая отщепление
фосфорной кислоты от АТФ:
АТФ + Н2О
= АДФ + H3PO4
3.7.Гидролизуют
С-С-связи.
Например, оксалоацетаза,
ускоряющая гидролитическое расщепление
оксалоацетата до оксалата и ацетата.
Лиазы [3]
Лиазы катализируют
отщепление от субстратов тех или иных
групп без участия воды или фосфорной
кислоты. В результате действия этих
ферментов образуются двойные связи.
Лиазы могут участвовать в реакциях
присоединении отдельных групп или
радикалов к двойным связям (воды, СО2,
NH3).
Класс лиаз делят
на семь подклассов. Это деление основано
за том, каков тип подвергающейся разрыву
связи между отщепляемой группой и
остатком молекулы.
Систематические
названия ферментов этого класса образуют
из названия субстрата, затем указывают
отщепляемую группу и через дефис
добавляют слово лиаза.
4.1. С-С-Лиазы
(углерод-углерод-лиазы)
— многочисленные ферменты, катализирующие
отщепление или присоединение углекислоты.
Отщепление углекислоты от органических
соединений часто происходит в процессах
обмена веществ в растениях. Ферменты
этого типа носят название карбоксилаз
или декарбоксилаз.
Пируватдекарбоксилаза
(4.1.1.1) катализирует отщепление СО2
от пировиноградной кислоты:

Кроме пировиноградной
кислоты, она может катализировать
отщепление углекислоты и от других
а-кетокислот с образованием соответствующих
альдегидов. Пируватдекарбоксилаза
— двухкомпонентный фермент. В состав
его активной группы входит производное
витамина В1
— тиаминпирофосфат.
В процессе фотосинтеза
важнейшую роль играет фермент
рибулезодифосфаткарбоксилаза
(4.1.1.39), катализирующий реакцию:

ФЕП-карбоксилаза
(4.1.4.31)
Глутаматдекарбоксилаза
(4.1.1.15)
участвует в декорбоксилировании
глутаминовой кислоты.
Аспарат-1-декарбоксилаза
отщепляет СО2
от аспарагиновой кислоты с образованием
новой аминокислоты в-аланина:
![]()
Это также
двухкомпонентный фермент, активная
группа которого представляет
фосфорилированное производное витамина
В6.
Декарбоксилазы
аминокислот
(4. 1.1 ) участвуют в отщеплении углекислоты
от аминокислот. В результате образуются
соответствующие амины:

При декарбоксилировании
АК лизина
лизиндекарбоксилазой
(4.1.1.18) образуется амин кадаверин
 ,
,
а при декарбоксилировании орнитина
образуется петресцин
 .
.
Оба вещества являются токсичными
(трупные яды).
Фруктозодифосфатальдолаза,
или альдолаза
(4.1.2.13) расщепляет шестиуглеродный
фруктозо-1,6-дифосфат на две триозы —
3-фосфоглицериновый альдегид и
фосфодиоксиацетон (фотосинтез, дыхание,
спиртовое бражение):

Лиазы кетокислот
(4. 1 .3) катализируют расщепление и
образование различных кетокислот. Среди
ферментов этого подкласса важное
значение имеет цитрат-синтаза
(4.1.3.7), с участием которой происходит
образование лимонной кислоты в цикле
ди- и трикарбоновых кислот:

4.2. С-О-Лиазы
(углерод-кислород-лиазы), или гидролиазы,
— многочисленная группа ферментов,
которая катализирует отщепление от
субстрата водорода и гидроксила,
образующих затем воду.
Карбонат-гидро-лиаза
или угольная
ангидраза
(4.2.1.1) расщеплфет угольную кислоту на
СО2
и воду: Н2СО3
СО2 +
Н2О
К этой группе
ферментов относится фумарат-гидратаза
(4.2.1.2) отщепляющая воду от яблочной
кислоты с образованием фумаровой
кислоты:
![]()
Фосфопируват-гидротаза
(4.2.1.11) или енолаза
участвует
в превращении 2-ФГК в 2-ФЕПК при гликолизе
и спиртовом бражении.
Цитрат(изоцитрат)
— гидро-лиазы (4.2.1.3)
катализирует взаимное обратимое
превращение лимонной, изолимонной и
цис-аканитовой кислот:
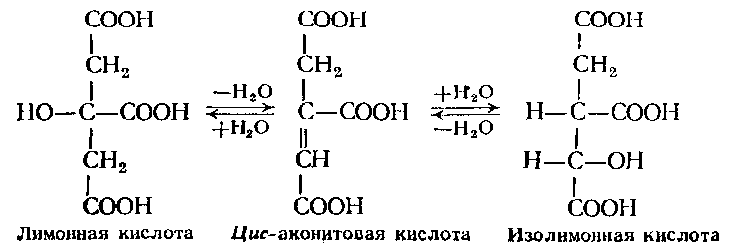
4.3. С-N-Лиазы.
Катализируют присоединение и отщепление
азотистых соединений.
Фермент
аспартат-аммиак-лиаза
(4.3.1.1) или аспартаза
катализирует взаимное превращение
аспарагиновой и фумаровой кислот:

Изомеразы [4]
Изомеразы катализируют
превращения органических соединений
в их изомеры. Изомеризация происходит
вследствие внутримолекулярного
перемещения атомов, атомных группировок,
остатков фосфорной кислоты, различных
радикалов.
Систематическое
название ферментов этого класса образуют
по схеме: субстрат-тип реакции
изомеризации-изомераза. При
внутримолекулярном переносе групп
ферменты имеют тривальные названия
мутазы,
а в реакциях инверсии групп у хиральных
центров — рацемазы
и эпимеразы.
В 5-м классе выделено
шесть подклассов.
5.1. Рацемазы
и эпимеразы.
Рецемазы катализируют взаимные
превращения D-
и L-изомеров,
эпимеразы — реакции эпимеризации,
заключающиеся в изменении взаимного
расположения атомов водорода и гидроксила
у одного из ассиметричных атомов
углерода. Одним из ферментов этого
подкласса является рибулозо-5-фосфат-3-эпимераза
(5.1.3.1), катализирующая превращение
ксилулозо-5-фосфата в рибулезо-5-фосфат:

ксилоза рибулоза
5.2.Цис-транс-изомеразы.
5.3. Внутримолекулярные
оксидоредуктазы. Эти
ферменты катализируют взаимное
превращение альдоз и кетоз, котонных и
енольных групп, перемещают С=С-связи. К
ним относятся триозофосфат-изомераза
(превращение ФГА в ФДОА),
 глюкозофосфат-изомераза
глюкозофосфат-изомераза
(превращение глюкозо-6-фосфата во
фруктозо-6-фосфат).

Известно более 20
ферментов, катализирующих превращение
альдоз и кетоз.
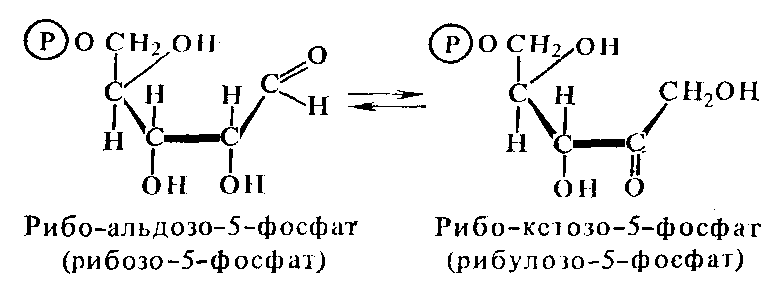
5.4. Внутримолекулярные
трансферазы
— осуществляют внутримолекулярный
перенос ацильных, фосфорильных и др.
групп. Примером фермента этого подкласса
является D-фосфоглицерат-2,3—фосфомутаза,
катализирующая превращение 3-ФГК
(3-фосфо-D-глицерат)
в 2-ФГК (2-фосфо-D-глицерат):
Лигазы (синтетазы)
[6].
Систематическое
название образуется по схеме: А:В лигаза
(образующая АДФ),
где А и В —
соединяющиеся молекулы, в скобках
указывается продукт расщепления
нуклеозидтрифосфата, который использовался
в данной реакции как источник энергии.
Весь класс делится
на пять подклассов в зависимости от
того, между какими атомами образуются
связи.
6.1. Образуют
С-О-связи.
Такой тип связи возникает при действии
аминоацил-тРНК-синтетаз,
активирующих аминокислоты и передающих
их на т-РНК.
6.2. Образуют
С-S-связи.
Ферменты этого подкласса катализируют
присоединение различных кислотных
остатков от КоА.
Ацетил-
КоА-синтетаза
катализирует реакцию: АТФ + CH3COOH
+ HS-КоА
= АМФ + H3PO4
+ CH3COO-KoA.
6.3. Образуют
С-N-связи.
Эти ферменты участвуют, например, в
синтезе различных амидов: аспарагинсинтетаза,
глутаминсинтетаза.
Аспарагиновая
кислота + АТФ = аспарагин + АДФ + H3PO4
6.4. Образуют
С-С-связи.
Сюда относятся карбоксилазы.
Эти Ф при участии АТФ катализируют
присоединение СО2
к различным органическим кислотам, т.е.
реакции удлиннения углеродной цепочки.
Эти ферменты содержат витамин
биотин. Одним
из представителе таких ферментов
является пируваткарбоксилаза
(6.4.1.1):
ПВК + СО2
+ АТФ + Н2О
ЩУК + АДФ + Н3РО4
Существуют шесть классов ферментов
В 1961 г в Москве V Международный биохимический союз принял современную классификацию ферментов. В соответствии с этой классификацией все ферменты делятся:
- на классы – по типу катализируемой реакции,
- каждый класс подразделяется на подклассы – по природе атакуемой химической группы,
- подклассы делятся на подподклассы – по характеру атакуемой связи или по природе акцептора.
Выделяют 6 классов ферментов:
- I класс – Оксидоредуктазы (подробно)
- II класс – Трансферазы (подробно)
- III класс – Гидролазы (подробно)
- IV класс – Лиазы (подробно)
- V класс – Изомеразы (подробно)
- VI класс – Лигазы (подробно)
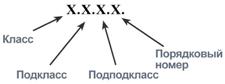
Каждому ферменту присвоен четырехзначный классификационный номер, включающий класс, подкласс, подподкласс и порядковый номер в подподклассе.
Например, алкогольдегидрогеназа имеет номер КФ 1.1.1.1. – это оксидоредуктаза, действует на ОН-группу донора с НАД в качестве акцептора с первым порядковым номером в своем подподклассе; лактатдегидрогеназа – КФ 1.1.1.27, действует на ОН-группу донора с НАД в качестве акцептора с порядковым номером 27 в своем подподклассе
Чтобы дать ферменту название существует два способа:
1. Систематическое название – согласно современной классификации. Часто такое название длинно и сложно для использования, поэтому как производное систематического названия у многих ферментов имеется одно или несколько рабочих названий.
2. Тривиальное название – название, сложившееся исторически, например, пепсин, трипсин, папаин, бромелайн, химозин. Для некоторых ферментов (чаще для гидролаз) к названию субстрата добавляется окончание «-аза» – инвертаза, уреаза, амилаза, лактаза, липаза. Тем не менее и у таких ферментов имеется систематическое название.
В 2018 году было предложено ввести новый, седьмой класс ферментов – Транслоказы (подробнее). Причиной стало то, что ни один из принятых классов не может описать группу ферментов, катализирующих движение ионов или молекул через мембраны, или их разделение внутри мембран. Некоторые из этих ферментов связаны с гидролизом АТФ и ранее классифицировались как АТФазы (ЕС 3.6.3.х), хотя гидролитическая реакция не является их основной функцией.
Шифр КФ («классификация ферментов»), или код фермента — классификационный номер фермента (b) по международной иерархической классификации. Принятая система классифицирует ферменты по группам и индексирует индивидуальные ферменты, что важно для стандартизации исследований.
Классификация ферментов основана и периодически обновляется Комиссией по ферментам (англ. (b) Enzyme commission, отсюда термин «EC number», принятый в англоязычной литературе) при Международном союзе биохимии и молекулярной биологии (b) . Каждый шифр КФ ассоциирован также с рекомендованным названием соответствующего фермента. Классифицировано более 3500 ферментов.
Принцип классификации
Классификация ферментов учитывает реакционную и субстратную специфичности ферментов, а не их белковую (b) структуру. Шифр КФ определяет химическую реакцию (b) , катализируемую ферментом. По этой причине аналогичные ферменты (иногда десятки) из различных организмов имеют один КФ, несмотря на структурные различия.
Иногда различные ферменты одного организма имеют один КФ. Например, панкреатическая липаза (b) и печёночная липаза (b) обе принадлежат к КФ 3.1.1.3, поскольку катализируют одну и ту же химическую реакцию (гидролиз (b) эфирной связи (b) в триглицериде (b) ), хотя первый фермент является ферментом пищеварения (b) и работает в кишечнике (b) , а второй относится к ферментам обмена липопротеинов (b) в крови (b) .
Существует база данных UniProt (b) [1] , которая идентифицирует каждый белок по его первичной последовательности. Обе базы данных дополняют друг друга.
Формат шифра
Каждый классификационный номер содержит сокращение КФ и последовательность из четырёх чисел, разделённых точками, и составляется по определенному принципу. Каждое последующее число представляет собой всё более и более уточняющую классификацию фермента. Так как база данных постоянно обновляется, коды могут меняться, причём коды некоторых уровней могут оставаться незаполненными.
Класс
- Код первого уровня (первое число, может быть от 1 до 7) указывает номер одного из семи главных классов ферментов (см. таблицу Коды первого уровня).
Подкласс
- Код второго уровня (второе число) означает подкласс, характеризующий основные виды субстратов (b) , участвующих в данном типе химических реакций. Например, у трансфераз (b) вторая цифра указывает на природу той группы, которая подвергается переносу, у гидролаз (b) — на тип гидролизуемой связи и т. д.
Под-подкласс
- Код третьего уровня (третье число) определяет более частные подгруппы (под-подклассы), отличающиеся природой химических соединений доноров (b) или акцепторов (b) , участвующих в данной подгруппе реакций. У гидролаз (b) , например, эта цифра уточняет тип гидролизуемой связи, а у лиаз (b) — тип отщепляемой группы и т. д. Первые 3 числа шифра точно определяют тип фермента.
Код четвёртого уровня
Наконец, все ферменты, относящиеся к данному под-подклассу, получают свой порядковый номер (четвёртое число в шифре).
- Например, глюкозоксидазе (b) присвоен шифр КФ 1.1.3.4, что означает:
- КФ 1 — Оксидоредуктазы (b)
- КФ 1.1 — Алкогольоксидоредуктазы (b)
- КФ 1.1.3 — Оксидоредуктазы, окисляющие группу CH-OH и восстанавливающие кислород (b) .
- КФ 1.1.3.4 — Оксидоредуктазы, окисляющие глюкозу (b) в присутствии кислорода (b) . Всего известно 3 глюкозоксидазы из разных организмов.
Коды первого уровня
| Класс | Катализируемая реакция | Тип реакции | Примеры |
|---|---|---|---|
| КФ 1 Оксидоредуктазы (b) | Окислительно-восстановительные реакции. Перенос атомов H и O или электронов (b) от одного субстрата на другой | AH + B → A + BH (восстановление) A + O → AO (окисление) | дегидрогеназа (b) , оксидаза (b) |
| КФ 2 Трансферазы (b) | Перенос функциональной группы (b) от одного субстрата на другой. Это может быть метильная, ацильная, фосфатная группа или аминогруппа. | AB + C → A + BC | аминотрансфераза (b) , киназы (b) |
| КФ 3 Гидролазы (b) | Образование двух продуктов из одного субстрата в результате гидролиза (b) . | AB + H2O → AOH + BH | липаза (b) , амилаза (b) , протеаза (b) , фосфатаза (b) |
| КФ 4 Лиазы (b) (синтазы) | Негидролитическое добавление или удаление группы к или от субстрата. Образование C-C, C-N, C-O или C-S связи. | RCOCOOH → RCOH + CO2 | карбоксилазы (b) |
| КФ 5 Изомеразы (b) | Внутримолекулярная перестановка, то есть изомеризация (b) молекулы субстрата. | AB → BA | изомеразы (b) , мутазы (b) и др. |
| КФ 6 Лигазы (b) (синтетазы) | Соединение двух молекул в результате синтеза новой C-O, C-S, C-N или C-C связи (b) , сопряжённое с одновременным гидролизом АТФ (b) . | X + Y+ ATP → XY + ADP + Pi | синтетазы (b) |
| КФ 7 Транслоказы (b) | Перенос ионов или молекул через мембраны или их разделение в мембранах. | мембранные транспортные белки[2] |
История
Схема номенклатуры ферментов была впервые разработана в 1955 году (b) , когда Международный конгресс биохимии в Брюсселе (b) учредил Комиссию по ферментам (Enzyme Commission). Первая версия номенклатуры появилась в 1961 году (b) и включала около 900 ферментов, в версии 1978 года (b) было более 2000 ферментов. Версия 1995 года (b) содержит более 3500 ферментов.
См. также
- Ферменты (b)
Примечания
- ↑ ENZYME (Enzyme nomenclature database). ExPASy. Дата обращения: 14 марта 2006. Архивировано из оригинала 18 февраля 2006 года.
- ↑ Международный союз биохимии и молекулярной биологии. Новый класс ферментов: транслоказы.. IUBMB NEWS (август 2018).
Ссылки
- Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (NC-IUBMB), Официальный сайт
Ферменты (b) | |
|---|---|
| Активность |
|
| Регуляция |
|
| Классификация |
|
| Типы |
|